"The tradition in biology is to try to understand intracellular communication from the bottom up," says Adam Arkin, a member of the Department of Computational and Theoretical Biology in the Physical Biosciences Division. "This involves dissecting cellular function to the point that every molecular reaction in a biochemical pathway is known - leading to reaction networks so dense, with so many interconnections, that a flowchart of just the most important pathways in a simple cell covers a wall."
Arkin, who is also an assistant professor in Bioengineering and Chemistry at UC Berkeley, says "it's difficult for biologists to make sense of what happens when reactions in such a dense and tangled network are modified even in a bacteria-much less to predict, in human cells, how particular mutations lead to disease, or how particular pharmaceuticals will affect an individual."
That's why he and his colleagues "want to design tools for understanding
cellular biochemical networks which are similar to the tools electronics
engineers use to design circuits," he says. "For all their complexity,
there are interesting ways in which the circuitry of a living cell resembles
electronic circuitry."
A living cell is not (quite) a digital computer
 |
||
 |
||
| Cell circuitry is analogous to electronic circuitry, but there's a lot more of it. The schematic above shows the reactions in E. coli metabolism by which the cell transforms matter. If regulatory interactions were added, the panel would be so dense with detail it would be black. | ||
At first glance the analogy between electronic circuits and living cells seems far-fetched. Diverse atoms and molecules, not indistinguishable electrons, are the cell's currency-units as simple as elemental ions, as specific as enzymes, or as complex as long strands of dna or rna, each with its own unique set of sources and targets.
And while an electrical circuit consists of an arrangement of physical parts, the "parts" of a biochemical network are mostly chemical reactions themselves-not entities but processes, whose existence depends on the presence of the right chemicals at the right place, at the right time, and in the right amounts.
The number of possible states in even elementary chemical "devices" is usually much greater than two; where large numbers of molecules are involved, the ratio of chemical concentrations may change virtually continuously. Still, the outcome of a given biochemical process is often binary, like an on/off switch.
For example, the amino acid glycine is produced from 3-phosphoglycerate by a series of enzyme-catalyzed reactions. If the supply of phosphoglycerate runs out, the reaction shuts down-but more often it shuts down when too much glycine inhibits the first reaction step; in either case the result is "off."
Arkin and his collaborators have identified several classes of chemical devices which function like basic units of electronic circuits, such as "voltage dividers" that can divide or amplify chemical signals and "frequency filters" that respond only to particular timing patterns in chemical concentrations. These sets of reactions are "wired" in series or parallel to form regulatory feedback loops, binary switches, oscillators, and even-in a resonant metaphor from biology to electronics and back again-computer-like "neural nets" that calculate the cell's needs and adapt to changing conditions.
Reactions involving large numbers of molecules are capable of deterministic outcomes, but when important functions make use of very small numbers of molecules, the outcomes are not completely assured; rather, they are stochastic.
Noisy signals and missing clocks
 |
|
 |
|
 |
|
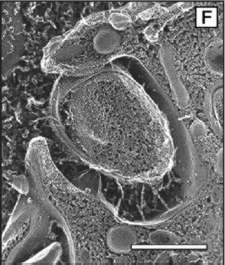
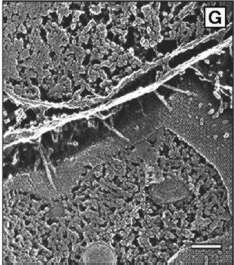
| E. coli cells are normally free-floating, but they can grow pili ("hairs") and attach themselves to tissue, such as the lining of the urethra. Adam Arkin and his colleagues found the switch that controls expression of pili; it operates randomly. | |
When a crucial signal consists of less than a hundred molecules, disruptive jostling among them, known as thermal noise, is high; how the reaction proceeds and which of the possible signals gets sent is a random affair. The process moves by fits and starts, and the time to complete it varies-although once underway, buildup of chemical concentrations is usually self-reinforcing.One way electronic circuit designers improve the signal-to-noise ratio is through signal synchronization. But unlike most digital circuits, biochemical networks (with a few exceptions) have no clock. The timing of even seemingly regular oscillations such as cell growth and division varies widely and can hang up, waiting until swarms of internal processes are completed and external conditions, like nutrient supply, are right.
"Synchronous design cuts noise by ignoring signals that arrive at the
wrong time-but asynchronous systems also have advantages, which even chip
designers are beginning to appreciate," says Arkin. "An asynchronous circuit
can speed up or slow down in response to changing conditions like temperature.
There's no circuitry wasted maintaining synchrony, and if a new version
of a component is placed in the system, the network can adapt to the change.
Power requirements are adjustable and generally lower." Arkin points out
that "these are just the sorts of advantages that favor evolution by natural
selection."
Survival through stochastic logic
Arkin and his colleagues, including Harley McAdams and John Ross of Stanford University, have analyzed the stochastic logic of gene regulation in several cell systems, including well-known, long-investigated genetic switches in viruses and bacteria.
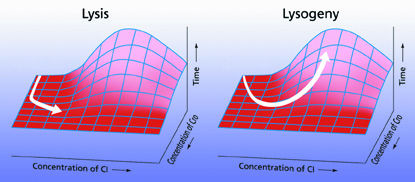
When a bacteriophage ("bacteria eater") virus injects its own dna into a microorganism such as Escherichia coli, the host cell apparatus rapidly expresses the program on the viral dna that decides whether or not to kill the host immediately. Under conditions that are less than optimal for replication, the phage may actually confer immunity to further infection upon the host (lysogeny). But if conditions are good, the virus produces so many copies of itself that the cell walls burst-a state known as lysis-and the infection spreads.
Two independently produced regulatory proteins compete to control whether the invading genes will remain quiescent or be expressed. Because of inescapable thermal noise, the outcome in any given case is random, and the proportion of the population in either state changes according to conditions such as cell nutrition and the number of invading particles per cell.
Arkin and his colleagues have found that the underlying stochastic mechanisms of the lysis-lysogeny decision circuit, which involves five genes and four promoter proteins, depend entirely upon the chance timing and concentrations of bursts of competing proteins that act to reinforce or inhibit one another.
"Thermal fluctuation at the molecular level makes for diversity in cells that start out under identical conditions," says Arkin. "The phage actually makes use of noise as a survival mechanism: sometimes it pays to multiply and infect as many hosts as possible, sometimes it pays to lie low. Either way, the viral population is prepared to cope with changing conditions."
E. coli itself adapts a comparable strategy to increase its survival chances. In humans, most E. coli live benign and even useful lives in the bowel. The bacteria can invade new territory, however, by sprouting hairlike filaments known as pili and anchoring themselves to tissues such as the lining of the urethra-where they resist being washed away and can cause painful and dangerous infections.
Immobility is a perilous advantage. Pili make it hard for bacterial cells to divide and multiply, and they can also trigger the body's immune system; their proteins make fine binding sites for antibodies. In any population of uropathic E. coli, only a few at a time display pili.
 |
|
| Two regulatory proteins control whether an infecting phage's replication genes will be expressed in E. coli-or if genes for integrating the virus into the host genome, and repressing other phage, will be produced instead. If the concentration of cI protein remains low, the replication genes are expressed (lysis); if cI concentration increases, the system regulates itself to prevent their expression (lysogeny). The concentration is a matter of chance. | |
"What causes an individual cell to express pili or not is a dna inversion switch," says Arkin, "in which a protein clips the gene in such a way that it reads 'pili' in one direction and 'no pili' in the other." Arkin and his colleagues have been investigating how noise in the system leads to random differences in the rate at which individual bacteria switch pili on and off, under different environmental conditions-"which leads to significant differences in the proportion of hairy cells in a population, which affects its spread and colonization."
Biochemical switches such as the lysogeny/lysis decision in viruses,
governed by antagonistic feedback loops, and pili expression in bacteria,
randomly implemented by a dna inversion reaction, are found in many different
infectious microorganisms. Indeed, they are typical of developmental pathways
in general.
Simulating cell circuits
Arkin and his colleagues have investigated pathways in more complex cells, such as the calcium-ion oscillator that may control embryonic cell division in organisms such as sea urchins and frogs. Nevertheless, he is in no hurry to abandoned simple organisms. "The only way we can fully understand even a simple organism is by a formal mathematical framework, a rigorous and testable theory, linked to experimental results," he says. "What goes on in a single cell really is a big deal."
With testable predictions derived from theory and computer modeling, and with a growing database of chemical devices and circuit motifs, Arkin hopes to make the understanding of intracellular communication and control "a true engineering discipline. We've hardly begun, but we're getting the tools."
One of the primary tools he is working to develop is a computer-aided simulation and analysis program called bio/Spice-a name deliberately chosen to echo spice (for "Simulation Program for Integrated Circuit Evaluation"), developed at the University of California at Berkeley in the 1970s and widely used to analyze electronic circuits and predict their behavior.
With bio/Spice, Arkin hopes to create a way for biologists to understand and build models of living systems as easily as electronic-circuit designers lay out a chip. bio/Spice and other tools will "yield new insights and new biological principles," he says, and "aid in designing novel functions into cells."
If Arkin is right, both electronic and cellular circuitry are subsets of overarching circuit-design principles. He thinks it's possible that a basic understanding of intracellular communication "may even teach us how to design more robust electronic circuitry."
Meanwhile the focus is on fundamental biology. "The Department of Energy's national laboratories have the best data-production and analysis facilities in the world-from tools for decoding the genome, to determining protein structure, to analyzing the cellular environment and whole tissues," Arkin says, "These are the facilities-and the people-we need to ask the complex questions. And the opportunities are spectacular." - end -
| < Research Review | Top ^ | |